Developmental Stages in Human Embryos
Go to Stage: Intro 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
Stage 16
Page 187Approximately 11–14 mm
Approximately 37 postovulatory days
SUMMARY
External: the nasal pits are coming to face ventrally and to disappear from profile view; retinal pigment is becoming visible externally; pharyngeal arch 2 is more massive and more conspicuous, whereas arch 3 is receding from the surface; auricular hillocks are beginning to appear; the thigh, leg, and foot are becoming distinguishable.
Internal: foramen secundum is present in the heart; a definitive mesentery has appeared and intestinal rotation is commencing; the pelvis of die ureter is becoming bipartite; no longitudinal fissure is found yet between the cerebral hemispheres; the neurohypophysial evagination is developing in the more advanced embryos; the lens pit is D-shaped; thickenings for the semicircular ducts are appearing.
SIZE AND AGE
As more-advanced and larger embryos are considered, there appears to be an increasing range in variability of size. If, however, size be regarded from the standpoint of the percentage variation in the greatest length of the embryos, the size range in these groups is not greatly different from those at previous stages. This should be taken into account in considering both actual size differences and differences resulting from the condition and treatment of the embryos.
The greatest length is most frequently 8–11 mm, but about one-fifth of the embryos are around 7 mm and slightly fewer are 11–12 mm. The size of the chorion is very variable, and there is much overlapping with the preceding stage. The range is from 15 to 48 mm.
The age is believed to be approximately 37 postovulatory days. Specimens that may well belong to this stage have been recorded as the 38th day (Tandler, 1907) and the 40th day (v. Hayek, 1934).
EXTERNAL FORM
The general proportions and shape of the head of the embryos at this stage are illustrated in figures 16-1 to 16-5. It will be seen that these are fairly uniform, largely because of the brain, the main parts of which can be recognized in the intact embryo (O'Rahilly, Müller, and Bossy, 1982). The mesencephalic and thalamic regions are relatively longer and more prominent than they were in the preceding stage. It is in the details of special regions, however, that developmental progress can be most sharply identified, and in this group such distinctive characteristics are to be found in the nose, eye, hyoid arch, and especially in the limb buds.
The nasal areas made their appearance earlier as two bilaterally symmetrical, discrete, and separate plates. In the present stage they are still separate and discrete. When they acquired their elevated borders, in earlier specimens, the discs faced laterally and their concave floors could be seen in profile views of the embryo. In the less advanced members of stage 16 a part of the floor of the nasal pit can still be seen in profile views, but in more advanced members, because of the increased relative thickness of the surrounding tissues, the pits face ventrally and their floor is no longer visible. One sees only the prominent lip which now forms their lateral boundary. In all the specimens of the group the marginal fold (nasal wing) now overhangs the floor of the depression. It is further to be noted that, in the embryos belonging to the more advanced half of the group, a nasofrontal groove is making its appearance and marks the frontal border of the nose, being exaggerated in shrunken specimens.
The pigment in the external layer of the retina is now visible macroscopically in the intact specimen in the more advanced members of the group. It is best Page 188 seen in the fresh state or in specimens fixed in formalin.
As a consequence of the widening of the head, the two nasal pits turn ventrally and are brought relatively nearer to each other. Similarly the dorsal segments of the mandibular and hyoid arches are pressed laterally while their ventral ends are drawn medially and thereby partially disappear from the profile view. On that account these arches and the formation of the face must be studied in decapitated specimens and reconstructions. In the present stage the hyoid arch is not bent under toward the median plane as markedly as the mandibular arch, and consequently it is more conspicuous than the latter in profile views. It also appears more massive because of the superficial cell proliferations that constitute the precursors of the auricular hillocks. In stage 16 they can be seen on the hyoid arch in every specimen. These superficial masses are not to be confused with the primary grooves that subdivide the mandibular and hyoid masses as a whole into dorsal and ventral segments. Except in the less advanced members of the group, the third pharyngeal arch has receded from the surface and is covered over by its more bulky neighbor.
The cervical sinus, an indication of which may be detected in some embryos at stage 13, is clearly visible by stage 14. Its floor is formed by arches 3 and 4 (O’Rahilly and Müller, 1985). The cervical sinus is smaller by stage 16. Contrary to Frazer (1926), it would seem that pharyngeal arch 3 is no longer visible on the surface after stage 16. The operculum frequently shown in drawings is incorrect.
In the preceding stage, one could distinguish a rounded hand region as a discrete part of the upper limb bud, distinct from the arm and shoulder. In stage 16 the hand region has become differentiated into a central carpal part, around the edge of which there projects a thick crescentic flange, which is to form the digital plate. In transparent specimens one can usually see the marginal vein coursing along the rim of the latter. The lower limb bud, although bearing some resemblance to the upper bud, has its own characteristics from the outset. The two are never identical. They appear so at first, but that is only because in their simpler form there are fewer features to be compared. Closer study, however, reveals their respective individualities. In the present stage, three growth centers are visible in the lower limb bud, subdividing it into three principal regions. These are (1) a rostrolateral part, in which the femoral and obturator nerves of the lumbar plexus terminate, (2) a caudomedial part, in which the peroneal and tibial nerves of the sacral plexus terminate, and (3) the foot region, to the base of which a part of the tibial nerve already extends. Roughly these represent the thigh, leg, and foot. The pelvis is not yet clearly set off from the thigh. Outline forms of the lower limb bud, as found in stages 15–17, are shown in figure 16-4.
Spinal ganglia are evident on the surface throughout the whole length of the trunk. Somites can be seen distinctly in the caudal part of the trunk.
Face
The oral opening of embryos of stage 16 is still equipped with merely a future lower jaw. The increasing breadth of the brain determines the considerable distance between the two nasal pits, because they lie ventrolateral to it. The roof of the mouth (stomodeum) slopes ventrally between the nasal pits as a broad, smooth surface, curving dorsally over the rostral end of the head, with no interruption to mark the location of a future upper jaw. Sketches of this region in two embryos belonging to this stage are shown in figure 16-5. The pertinent details should be compared with those of embryos of the next stage, shown in figure 17-3.
The rnesenchyme will play an important part in the formation of the structures of the skull and face. The mesenchyme of the head is by no means a homogeneous tissue. Instead, it includes cell groups that come from more than one source and that have different potentialities. The group best known consists of the cells that emerge as a sheet or layer from the primitive streak. From these cells are derived, among other things, the somites (trunk muscles, axial skeleton, and meninges). Also some of them differentiate into angioblasts and give origin to the blood vascular system. Another important specialization of the mesenchyme results in a coalescence of its larger spaces into channels communicating with the exocoelom, producing an irrigation system, seen at the height of its development in stage 11. The surface cells of the coelom, under the influence of the regions in which they lie, respond by active proliferation and supply the visceral mesenchyme that gives origin to the vascularized muscular and connective-tissue coats of the viscera, whereas the residual surface cells become permanent serous membranes and serve as the pericardium, pleura, and peritoneum. These mesenchymal components, of primitive-streak ancestry, can be followed without much difficulty.
Page 189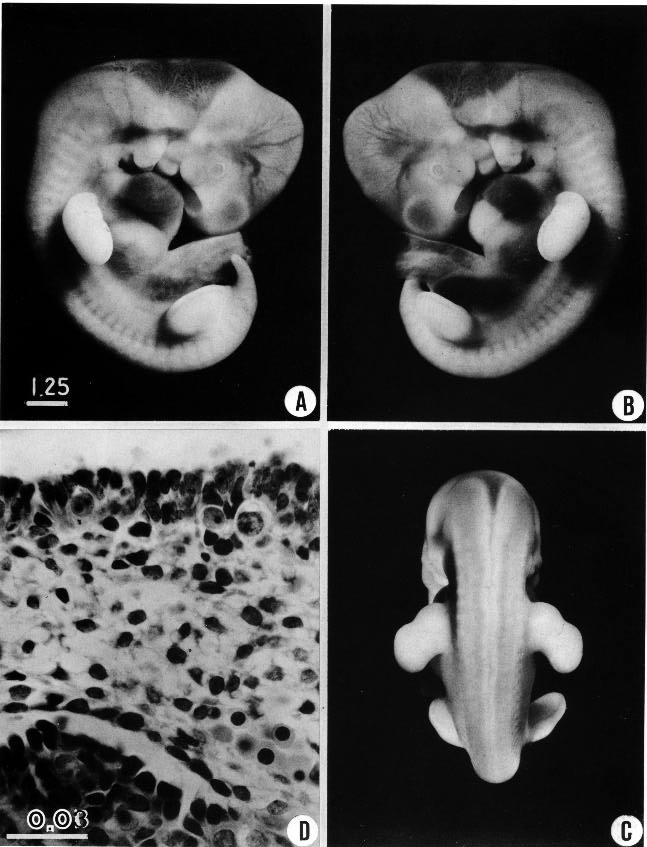
Fig. 16-1. (A-C) Three views showing particularly well the characteristics of a formalin-fixed specimen typical of the less advanced half of stage 16 (No. 8098). The limb buds are thicker than in the more advanced members of the preceding stage, and the marginal vein is now distinct in the lower limb bud.
(D) The gonadal region of the same embryo (section 10-5-4), showing proliferation and beginning invasion of the coelomic epithelium. Also shown is the early presence of primordial germ cells, three of which can be seen. The coelomic cells tend to cluster immediately around them. The larger pan of the picture consists of the reticulum of the mesonephros, with early angioblasts and a part of a glomerulus.
Page 190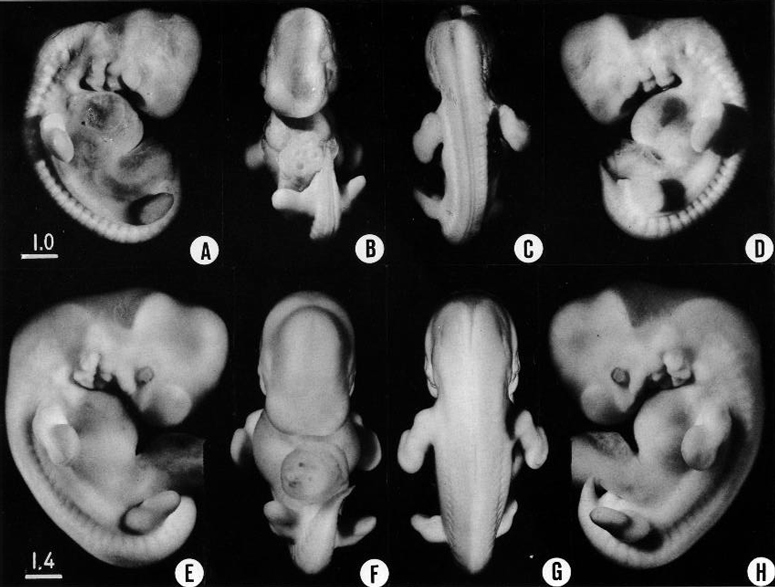
Fig. 16-2. Two embryos: one of the least advanced and one of the most advanced members of stage 16. The differences are exaggerated by the fact that the first specimen was fixed in formalin and then became shrunken. The examples illustrate the uncertainty of depending entirely on external form for alignment of embryos in their developmental order. Both specimens qualify for this stage on the basis of their internal structure. A and C show shrinkage and sinking in of the roof of the fourth ventricle. The distinctness of the ganglionic and somitic masses is also an effect of the shrinkage. The auricular hillocks and the limb buds are found to provide the most reliable of the surface characteristics for this period of development. (A–D) No. 6054 (E–H) No. 8112.
Page 191
Fig. 16-3. These embryos have been sectioned serially and in their microscopic structure qualify as members of stage 16. The differences between them are for the most part accounted for by the factors of fixation, shrinkage, manner of illumination, and original condition of the specimen. The otic vesicle, where it can be seen, the auricular hillocks, the lateral nasal wing, the disappearing third pharyngeal arch, the distinct hand plate, and the rounded foot plate with its early marginal vein are characteristics common to all these specimens (A) No. 6750. (B) No. 7115. (C) No. 7804. (D) No. 6517. (E) No. 6511. (F) No. 6510. (G) No. 6514. (H) No. 8179. (I) No. 7629. A–G are at the same magnification, H and I are slightly less.
Page 192Especially in the head region, other mesenchymal cells are known (chiefly from experiments on the chick) to be derived from the neural crest. A third source, the prechordal plate, furnishes cells for the muscles of the orbit (Gilbert, 1957) and probably for the meninges (O'Rahilly and Müller, 1986b).
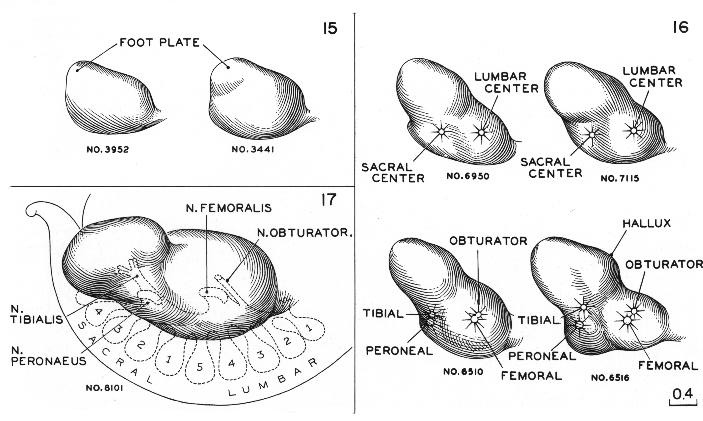
Fig. 16-4. Differentiation of the lower limb bud at stages 15–17. At stage 17 the primary divisions of the limb are clearly indicated by the nerve trunks, which branch in a characteristic pattern from the lumbosacral plexus. The femoral and obturator trunks lead to the thigh and, together with the condensed center surrounding them, are responsible for the bulging mass on the proximal part of the limb bud. Coincidently with the breaking apart of this mass into subcenters (muscles), the femoral and obturator trunks split into their respective branches. The trunks from the sacral plexus form the central core of the condensed mass of the caudal half of the limb bud, one trunk (N. tibialis) reaches distally to the plantar region of the foot plate. Thus the tibial and peroneal trunks delimit the primary parts of the limbs below the knee. This lumbosacral distribution might be expected from the fact that when the lower limb bud is first discernible at stage 13, its caudal half lies opposite the sacral nerve-ganglion masses, whereas its cranial half similarly lies opposite the lumbar ones. In stage 16, one can still identify the thigh and the leg by their neural relationships. The foot plate is already determined, and among the more advanced members of the group one can recognize the region of the hallux. Stage 15 marks the emergence of the foot plate. The stage is represented only by more-advanced members of the group. Among the less advanced members one finds less-discrete elevations of the limb bud.
Another characteristic of head mesenchyme is the prevalence of greater thickness of the skin ectoderm over its more condensed areas, suggesting increased functional activity. This is notable in the ventral parts of the head, particularly the mandibular and hyoid regions. The thick areas are part of the ectodermal ring (O'Rahilly and Müller, 1985). Such areas stand out in contrast with the loose reticular mesenchyme found over the dorsal parts of the head, where the skin ectoderm is thin, apparently merely adapting itself to the Page 193 expanding size of the underlying brain. Thick skin ectoderm (the apical ectodermal ridge) and marked proliferation of the mesenchyme with which it is in contact are likewise characteristic of the limb buds, and are an expression of interaction between the two tissues. The limb buds and the group of mesenchymal ridges that form at the sides of the mouth and pharynx around the termination of the large cranial nerves are the product of the same kind of developmental relation between the skin ectoderm and the underlying blastema, together with whatever the nerves may contribute.
In stage 16, between the thin skin ectoderm and the brain, there is found a sparse amount of loose embryonic connective tissue (the primary meninx) through which passes the network of capillaries and blood vessels supplying the brain wall and cranial nerves. Standing out in contrast are some fields where the mesenchymal cells by active proliferation have produced condensed areas, or detached foci of growth. They are found in the region of the eye, and surrounding the otic vesicle and the nasal plate. To some extent these condensations influence the surface form of the embryo, as seen in the lateral nasal process or wing.
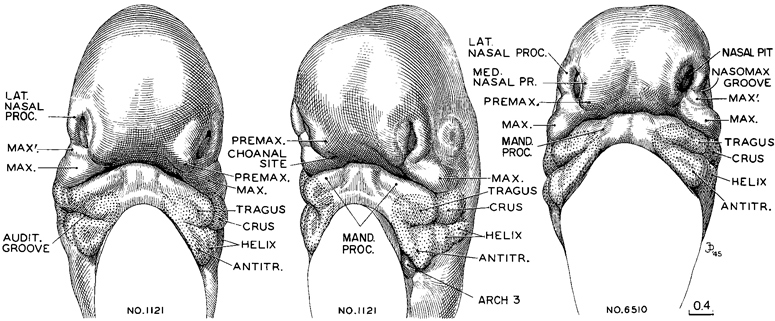
Fig. 16-5. Representative views of the nasal pits and oral region at stage 16. The two views of No. 1121 show a less advanced member of the group, whereas No. 6510 belongs among the more advanced ones. The right and left nasal pits show no evidence yet of uniting to form a nose. The interval between them consists of undifferentiated tissue extending from the roof of the stomodeum “forward” and “upward” over the frontal region of the head. This is sometimes spoken of as the “frontal process,” but if that term is to be applied at stage 16 one must be satisfied with an idea rather than with a recognizable structure. The growth centers that are to form the upper jaw are widely separated, like the nasal pits, and are present only as bilateral placed masses. These include the premaxillary (globular process of His) and maxillary centers, together with a characteristic subdivision of the latter (marked MAX'.).Though one cannot yet speak of an upper jaw, the future lower jaw is definitely marked off and hence has the distinction of being the first part of the facial equipment to become thus established. The auricular hillocks on the mandibular and hyoid arches are given labels with the pans of the external ear that they are to form. Drawings made by James F Didusch. Reconstructions made by Osborne O Heard. The drawings are all at the same scale.
In the more ventral parts of the head, mesenchymal condensation exhibits relative precocity. One can recognize a rather dense maxillary field that bulges on the surface as a prominent ridge (fig. 16-5, No. 1121). Under the surface this field blends with the adjacent condensations surrounding the nasal pit and eye of the same side. The right and left maxillary fields are, however, still widely separated. Between them is the broad region that is to become the roof of the mouth. The only surface markings of this region are two rounded, low elevations produced by incipient bilateral (premaxillary) condensations constituting the primordia of the primary palate. Apart from these modest representatives, there is lacking any future upper jaw. In many respects the ventral part of the cranium and the floor of the mouth are more advanced than its Page 194 sides and top. The nerves are larger, the epithelium is thicker and more specialized in its flange-like extensions, and the mesenchyme is more condensed and shows multiple foci of growth. There is a definite basis for the future lower jaw, and on the surface of the mandibular and hyoid arches are the condensations of the auricular hillocks, which proceed to form the specific parts of the external ear. These various changes take place before the advent of a cartilaginous skull.
CARDIOVASCULAR SYSTEM
Heart
By stage 16, “the flow of blood from the right ventricle to the sixth arches, and from the left ventricle to the fourth arches” is a fait accompli (de Vries and Saunders, 1962).
A typical series of sections through the heart at stage 16 is shown in figure 16-6. In such sections one can obtain an impression of the relative proportions and orientation of the different elements that make up the total heart. A conspicuous part is taken by the gelatinous reticulum, which thickly invests the primary cardiac tube everywhere except at the ventricular pouches. The latter consist of two trabeculated recesses opening from the ventral rim of the cardiac tube, as has been shown in three dimensions in figure 15-5. The cardiac mesenchyme (gelatinous reticulum) terminates abruptly on the aorta and pulmonary arch, where these become free of the heart. Proximal to this, the endothelium of the two is still found as a common aorticopulmonary trunk, although the two component regions can be identified. The semilunar valves are being formed. The site of the tricuspid and mitral valves consists of the mesenchyme that encircles the atrioventricular canal. It comprises rostroventral and caudodorsal cushions. The canal itself takes the form of a widening transverse slit, in the center of which the endothelium is disappearing, which results in the fusion of the endocardial cushions. This fusion, which occurs in stages 16–18 (McBride, Moore, and Hutchins, 1981), will produce two separate channels, the right and left atrioventricular canals.
For comparison, figure 15-5 shows how the single blood current of the primary cardiac tube becomes gradually subdivided into two diverse currents, the aortic and pulmonary, without any interruption, during the alterations, in the vital matter of delivery of umbilical and vitelline blood to the systemic vessels. Preceding the separation is a specialization of the primary cardiac tube in two definite regions along its rim. This is illustrated in the figure by the embryo representing stage 11. The primary tube is still a single channel from the atrioventricular junction to its outlet at the aortic sac. In the two separate regions that are to form the right and left ventricles, however, there is a trabeculated outgrowth of the endothelium. Although only endothelium is shown, the overlying myocardium also participates in this localized specialization, and in these areas sprouts from the outer myocardial shell extend inward and interdigitate with the endocardial trabeculae.
On passing to stage 13 in the same figure, the tube is seen to be more capacious and considerably longer in the region of the truncus arteriosus. This increased length may include the conus cordis. In stage 15, the now relatively more voluminous tube is in the process of separating into aortic and pulmonary channels. The separation is well under way in stage 16. The ventricular pouches invariably occur at the same sites, and, as one can see in stage 16, as they become larger their positions prove favorable for producing directional thrusts or currents toward the pulmonary and aortic outlets, respectively.
Details of the myocardium are shown in figure 16-7. Representative sections were taken of embryos of stages 14–17. One can see how the two trabeculated pouches (right and left ventricles) grow out separately from the primary cardiac tube. One also sees that when the pouches become larger and deeper, they coalesce and acquire a common party wall. The crest of the septum is laid down first and is added to below as the pouches become deeper. In other words, the interventricular septum grows from above downward, its crest being its oldest part.
Septum primum shows a club-shaped ending (the spina vestibuli of His), which extends between the atrioventricular cushions (Puerta Fonolla and Orts Llorca, 1978, fig. 3). Foramen primum is obliterated at stages 16 and 17 (McBride, Moore, and Hutchins, 1981). Other features of the heart at this time include fusion of the atrioventricular cushions with the interventricular septum (stages 16–18), and the appearance of lateral atrioventricular cushions and the conotruncal septum at stage 16. The atrioventricular node is said to be Page 195 identifiable (Yamauchi, 1965).
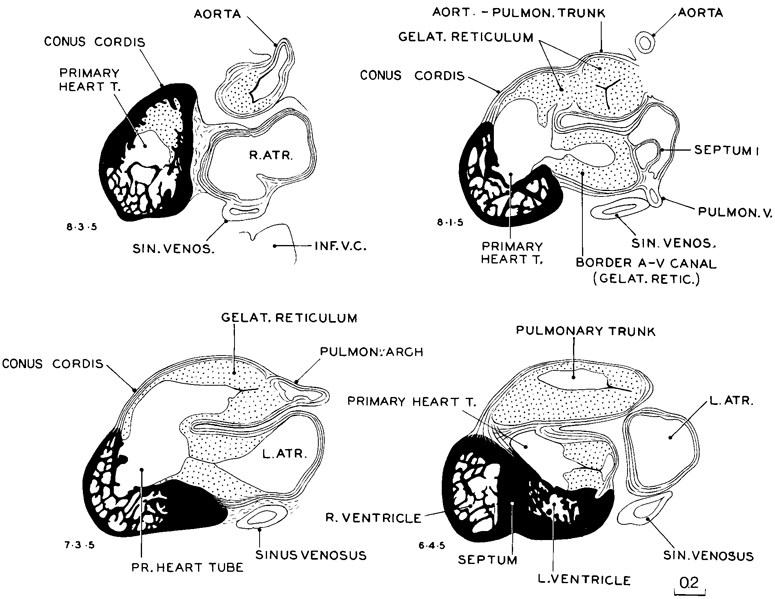
Fig. 16-6. Selected sagittal sections through the heart of No. 6511, typical of this stage. The primary cardiac tube, the wall of which is characterized by marked development of cardiac mesenchyme (gelatinous reticulum), constitutes a conspicuous part of the heart at this time. The ventricles opening off from it are shown in solid black.
A ventricle in general is considered to be “a chamber with one or more inlet portions” (Anderson, Wilkinson, and Becker, 1978), and each ventricle comprises three parts: an inlet adjacent to an atrioventricular orifice, a trabecular pouch, and an outflow tract (fig. 17-7). The details of the developing outflow tract in the human need further study. The truncus arteriosus, according to Los (1978), “participates in the development of the outflow tracts of the ventricles ... but not of the ascending aorta and the pulmonary trunk,” both of which are said to be derivatives of the aortic arches. Interventricular foramen 1 is never obliterated but is converted into part of the outflow tract of the left ventricle (Kramer, 1942).
The extracardiac portion of the aorta is situated to the right of and dorsal to the pulmonary trunk (e.g., Congdon, 1922, figs. 35 and 39, stages 16 and 17, respectively). According to Pexieder (1982), growth changes in the right ventricle and conus create an optical illusion of displacement of the conus to the left and of conal absorption. Pexieder maintains that the aorta is not transferred (i.e., displaced) to the left ventricle but rather bound to that ventricle by its infundibulum. The infundibulum is formed “behind” by the fused atrioventricular cushions and “in front” by the fused conal cushions, whereas its cavity represents interventricular foramen 1 (2 in Pexieder’s enumeration). Thus, in this view, the renversement d’alliance of the aorta from the right to the left ventricle is not Page 196 caused by displacement of the conus to the left, by reduction of the conoventricular flange, or by conal absorption.
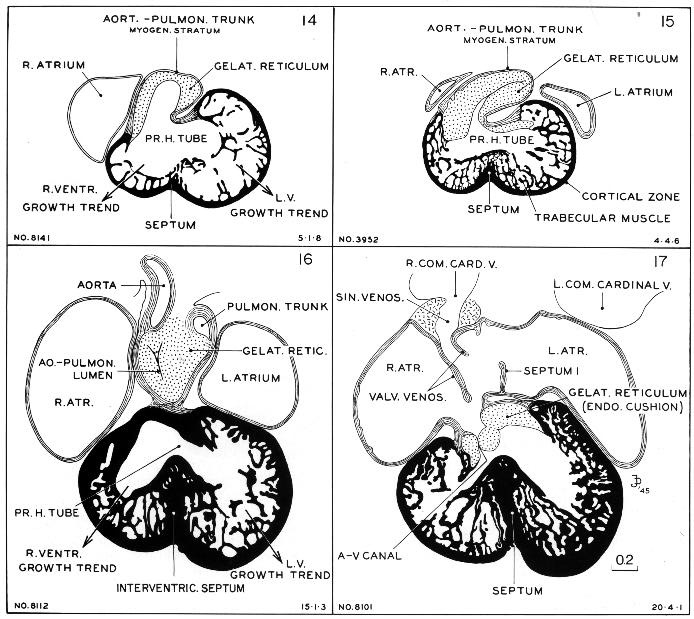
Fig. 16-7. The origin of the interventricular septum, as seen at stages 14–17. The sections of the first three stages are taken from embryos cut coronally and the fourth from one sectioned transversely. This makes them easily comparable. It will be seen that the two ventricles start as separate reticulated pouches, but as they expand their myocardial shells coalesce in the formation of a party wall. The crest of the latter is its oldest pan.
The aortic arches, in ventral view, appear to be more or less symmetrical at stages 11 and 12 but have begun to be asymmetrical at stage 16 and are definitely so from stage 17 onward (Congdon, 1922, figs. 1–16).
DIGESTIVE SYSTEM
The outlines of the epithelial canal for stages 15–17 (figs. 15-8, 16-8, and 17-8) were made for direct comparison of such features as relative length, diameters, flexures, and general form.
Page 197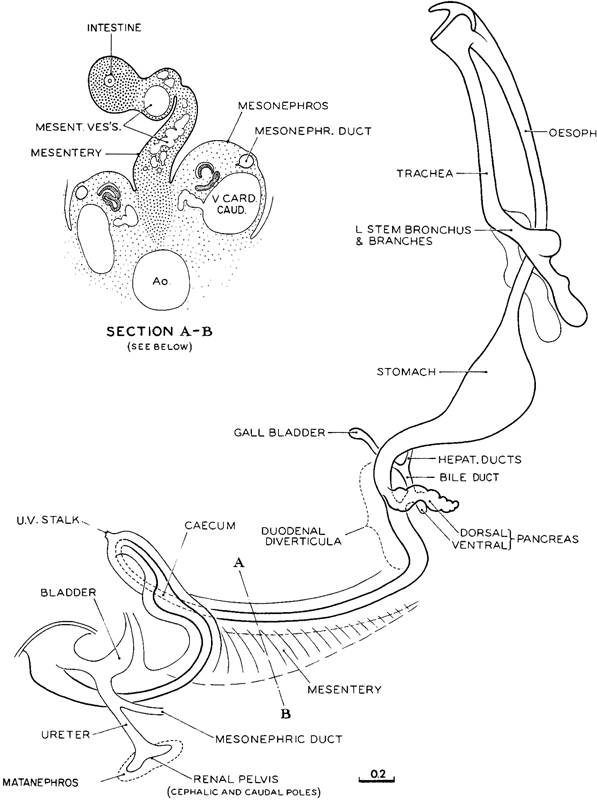
Fig. 16-8. Alimentary epithelium of No. 6510, a median member of stage 16. This drawing is to be compared with those of the next and preceding stages (figs. 15-8 and 17-8), shown at the same enlargement. Advancement in details of the separate pans of the system is evident, and it is to be noted that the trachea is now quite separate from the esophagus. There is now a definitive mesentery, the structure and attachment of which are shown schematically in the inset by a section (6-3-3) from No. 1121, cut in a favorable plane in the direction shown by line A–B. Drawing made by James F. Didusch. Reconstruction made by Osborne O. Heard.
A striking feature of the alimentary epithelium is the seeming leadership it exhibits in the delineation of the different parts of the system, and this applies also to such offspring as the pulmonary epithelium, biliary passages, and pancreas. These regions, which could be identified earlier, now stand out in advanced detail (fig. 16-8). It is to be noted particularly that one can now speak of a definitive mesentery, beginning near the caudal end of the duodenum and extending caudally to include the rostral half of the colon. It provides the framework for the primary intestinal loop. The section A–B in this figure shows schematically the structure of this mesentery and its terminus in the primordium of the gut wall. The mesenteric vessels are conspicuous. Here as elsewhere, distinct gut epithelium Page 198 is surrounded by condensed visceral mesenchyme showing early stages of angiogenesis. A distinction should be made between a definitive mesentery, such as is present at this stage, and the fanciful or theoretical mesenteries commonly pictured in textbooks.
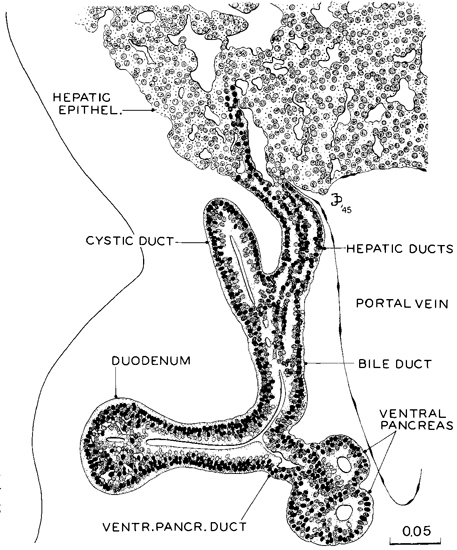
Fig. 16-9. Composite drawing of three successive sections showing the termination of the biliary ductal system at the duodenum, typical of stage 16 (No. 6511, sections 8-1-3 to 8-2-1). Only a small part of the cystic duct is shown. The abrupt termination of the hepatic ducts in the hepatic epithelium is characteristic. The bile duct serves as a stem maintaining the continuity between the hepatic epithelium and the duodenal epithelium, never interrupted from the time of the first outsprouting of the hepatic epithelium. Drawing by James F. Didusch.
Counterclockwise rotation of the gut commences. The originally proximal limb of the loop moves to the right, and the distal limb to the left.
In the duodenal region the epithelium is in the process of active proliferation, and with this there is a noticeable bulging of its contour. Two fusiform enlargements are thus found, separated by the ducts from the pancreas and the biliary system. A characteristic section through the duodenum at this time is shown in figure 16-9. This illustration indicates also the communications of the biliary system and the differentiation of the ducts. The ventral pancreas still empties by its own duct into the bile duct from which it was derived. The duodenal end of the dorsal pancreas is attached to the duodenum in front, and at a level nearer the pylorus than the bile duct. As can be seen in the figure, the growth of the hepatic ducts produces an intervening stem that isolates the hepatic epithelium from the lower biliary system. Examination of the extreme distal tip of the hepatic ducts, possible in the section illustrated, indicates that it is a differentiation in situ of the ductal epithelium (i.e., a conversion and proliferation of cells already present rather than a penetration by foreign cells). The section shown is a very early phase of the process. There are no discernible hepatic ducts at this time in any other part of the large hepatic mass. It has been claimed that, contrary to earlier accounts, the gall bladder is not connected with the hepatic mass (Lassau and Hureau, 1967). The lumen of the duodenum is probably continuous throughout (Boyden, Cope, and Bill, 1967).
Page 199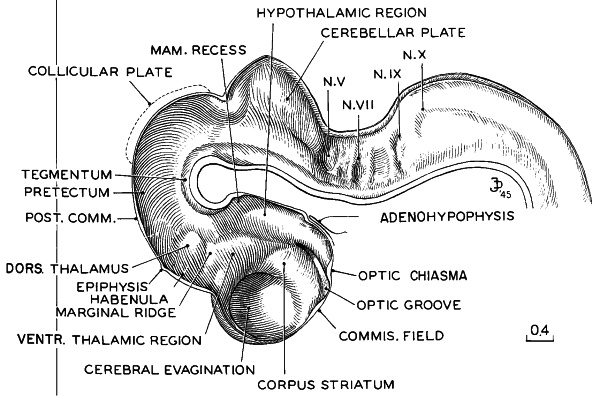
Fig. 16-10. Reconstruction of right half of brain of No. 6510, typical of stage 16. A marginal ridge now begins to mark off the dorsal from the ventral thalamus, the latter lying rostral to and below it. The tegmentum is extending caudally to become continuous with the basal plate of the hypoglossal region. A slight evagination of the infundibular floor in the median plane marks the beginning of the neurohypophysis. This figure is to be compared with the others of the brain series (figs. 15-9, 17-12, and 18-10), all of the same magnification. Drawing made by James F. Didusch. Reconstruction made by Osborne O. Heard.
RESPIRATORY SYSTEM
The pulmonary epithelium, which can be spoken of as a bronchial tree, is shown in figure 16-11. The right and left primary bronchi exhibit characteristic asymmetries. Each ends in a bulbous terminal growth center, and along the sides each gives off a characteristic type of elongating lateral branches. In the figure, three examples of development are shown, to cover the range that exists at this stage.
URINARY AND REPRODUCTIVE SYSTEMS
The metanephros, which is now reniform, is still sacral in level. The ureter is elongating and, in more advanced embryos, the pelvis of the ureter divides into rostral and caudal poles. The urorectal septum, the formation of which is disputed, is well marked.
The gonadal primordium is indicated by proliferation of the coelomic epithelium and the invasion of the underlying mesonephric stroma. The primordium of the paramesonephric duct is beginning to appear as an invagination of thickened coelomic epithelium over the mesonephros.
SKELETAL SYSTEM
It has been shown that, at stage 16, 36 elements for the future vertebral column are clearly delineated by the ganglia and myotomes (O'Rahilly and Müller, 1985, fig. 10). Two or three more elements have still to be added in order to complete the number in either the more advanced embryos of stage 16 or in stage 17. What Kunitomo (1918) designated the “longest tail” at stage 16 is nothing of the kind but is merely the caudal end of the embryo, which will develop into the coccygeal region.
Page 200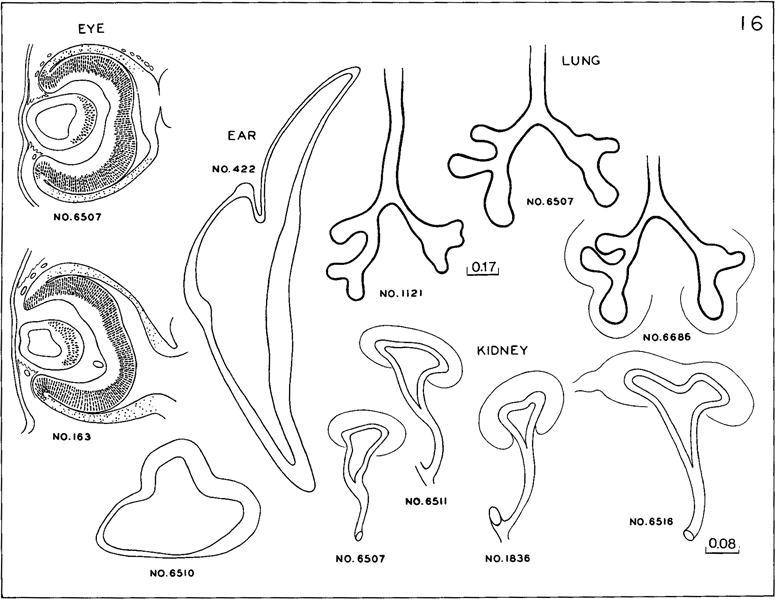
Fig. 16-11. Cluster of characteristics exhibited by each of the embryos assigned to stage 16. Retinal pigment is visible under medium magnification. The otic vesicle shows thinning of the wall that will later be absorbed. The primary bronchi have definite, elongating branches. The ureter is elongated and its pelvic expansion has an upper and a lower pole.
NERVOUS SYSTEM
The ascending fibers in the dorsal funiculus of the spinal cord reach the level of the medulla oblongata.
Rhombomeres are still present. The alar lamina is developing, and its upper part, the rhombic lip, is very dense. Cells of the general and special visceral efferent nuclei of the cranial nerves (5, 7, 9, 10, 11) are still migrating from medial to lateral. The reticular formation is better outlined than in stage 15. The growth of the cerebellar primordium in width and height deepens the groove between it and the mesencephalon. A well-pronounced isthmus rhombencephali is evident.
The mesencephalon still comprises two parts, and the button-like formation visible in stage 14 is still present in the roof between the two portions. The peripheral part of the trochlear nerve is present in all embryos.
The posterior commissure is beginning to develop. The dorsal and ventral thalami are separated by a groove adjacent to the marginal ridge (fig. 16-10). A neurohypophysial evagination is apparent in half of the embryos but is not very distinct. The medial (diencephalic) ridge of the corpus striatum is directly continuous with the lateral (telencephalic) ridge. The cerebral hemispheres Page 201 are bulging laterally, but no longitudinal fissure is present yet. The interventricular foramen is wide and is limited dorsally by the ventral thalamus. In the more advanced embryos the area epithelialis and sulcus limitans hippocampi become visible. The most developed telencephalic area is the amygdaloid part. Cajal-Retzius cells, with fibers tangential to the surface of the cerebral vesicles, are present. Olfactory fibers running from the nasal pit to the brain are developed in half of the embryos. The olfactory tubercle becomes apparent. Fibers of the medial forebrain bundle form.
The cells of the sympathetic nervous system form a solid trunk.
Eye
The growth of the lens body results in an approximately D-shaped lens cavity. Perilental blood vessels (tunica vasculosa lentis) are visible. Retinal pigment is shown in figure 18-12A.
Ear
The endolymphatic appendage is long. Thickenings in the wall of the main, or vestibular, portion of the otic vesicle presage the appearance of the semicircular ducts. The ventrocaudal portion of the otic vesicle, although not yet marked off, is the primordium of the cochlear pouch.
EMBRYOS OF STAGE 16 ALREADY DESCRIBED
No. BR, 9.75 mm G. L., 8.8 mm. Summarized by Tandler (1907), who, on the basis of the coital history, gave the age as the “38th day.” About stage 16.
9-mm embryo. The peripheral nervous system was described by Masy (1955) in this embryo of stage 16.
Huber No. 3, 10 mm. The nuclei of origin of the cranial nerves and the peripheral nervous system were described by Streeter (1908a,b). An advanced example of stage 16.
Wetzel (We) embryo, 10 mm. Discussed by v. Hayek (1934) in regard to age, which, based on the coital history, was given as the “40th day.” Probably about stage 16.
Carnegie No. 6516, 10.5 mm (corrected). Double ureters described by Wharton (1949).
No. H60, University of Missouri, 11 mm. Described briefly by Bonnot and Severs (1906). Probably belongs to stage 16.
Copyright © 1987 Carnegie Institution of Washington. Reproduced on ehd.org with permission.